เมื่อแสงมากเกินไป ใบพืชป้องกันตัวอย่างไร
 ภาพปกบทความ เมื่อแสงมากเกินไป ใบพืชป้องกันตัวอย่างไร
ภาพปกบทความ เมื่อแสงมากเกินไป ใบพืชป้องกันตัวอย่างไร
ภาพรวมของพืชภายใต้แสงจัด แสดงความสัมพันธ์ระหว่างใบ แสง ความร้อน ปากใบ ระบบราก และน้ำในดิน
กลไกการป้องกันตนเองของพืชภายใต้สภาวะความเครียดจากแสงส่วนเกิน
Mechanisms of Photoprotection under Excess Light Stress
แสงเป็นปัจจัยสำคัญที่ทำให้พืชสร้างอาหารได้
ใบพืชใช้พลังงานแสงเพื่อขับเคลื่อนกระบวนการสังเคราะห์ด้วยแสง (Photosynthesis) เปลี่ยนพลังงานแสงให้เป็นพลังงานเคมี และนำไปสู่การตรึงคาร์บอนไดออกไซด์ เพื่อสร้างน้ำตาลและสารอินทรีย์ที่พืชใช้ในการเติบโต
แต่ในธรรมชาติ แสงไม่ได้มีด้านเดียว
เมื่อแสงอยู่ในระดับที่ใบใช้ได้ทัน แสงคือพลังงาน แต่เมื่อแสงมากเกินกว่าที่ระบบใบจะนำไปใช้ได้ โดยเฉพาะในวันที่แดดแรง อากาศร้อน น้ำไม่พอ หรือปากใบปิดบ่อย พลังงานแสงส่วนเกินอาจกลายเป็นภาระต่อระบบสังเคราะห์แสงได้
สภาวะนี้เรียกว่า แสงส่วนเกิน (Excess light) หรือความเครียดจากแสง (Light stress)
ประเด็นสำคัญจึงไม่ใช่เพียงว่า “พืชได้รับแสงมากแค่ไหน” แต่ต้องดูว่า พืชสามารถใช้พลังงานแสงนั้น ระบายพลังงานส่วนเกิน และซ่อมแซมความเสียหายที่เกิดขึ้นได้ดีเพียงใด
บทความนี้ชวนดูว่า เมื่อแสงมากเกินไป ใบพืชป้องกันตัวเองอย่างไร ตั้งแต่การลดการรับแสง การระบายพลังงานส่วนเกิน การควบคุมอนุมูลอิสระ ไปจนถึงการซ่อมแซมระบบสังเคราะห์แสง และความเชื่อมโยงกับน้ำ อุณหภูมิ ธาตุอาหาร ฮอร์โมน และจุลินทรีย์ในระบบราก
1. แสงมากเกินไปคืออะไร
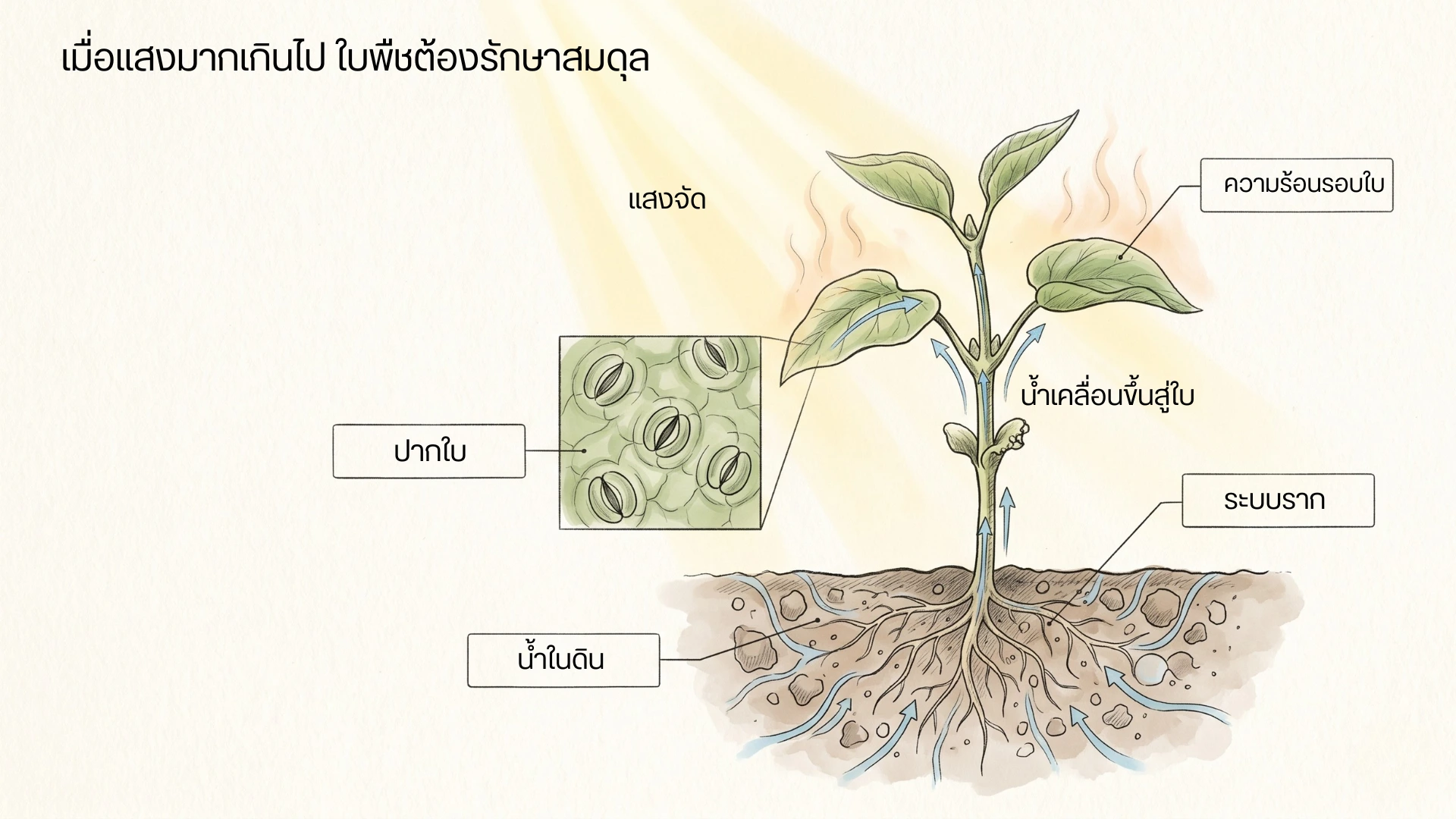 ต้นพืชภายใต้แสงแดดจัด แสดงความสัมพันธ์ระหว่างแสง ความร้อน ปากใบ ระบบราก น้ำในดิน และการเคลื่อนที่ของน้ำขึ้นสู่ใบ
ต้นพืชภายใต้แสงแดดจัด แสดงความสัมพันธ์ระหว่างแสง ความร้อน ปากใบ ระบบราก น้ำในดิน และการเคลื่อนที่ของน้ำขึ้นสู่ใบ
เมื่อแสงมากเกินกว่าที่ใบใช้ได้ทัน พืชต้องรักษาสมดุลระหว่างแสง ความร้อน น้ำ ปากใบ และระบบราก
แสงมากเกินไป ไม่ได้หมายถึงแสงที่มีความเข้มสูงเพียงอย่างเดียว
ในทางสรีรวิทยาพืช จุดสำคัญอยู่ที่ “แสงที่ใบรับเข้ามา มากกว่าความสามารถที่ใบจะนำไปใช้ในกระบวนการสังเคราะห์แสงได้ในขณะนั้น”
พลังงานแสงที่ใบรับเข้ามาต้องถูกส่งต่อไปยังระบบต่าง ๆ ภายในคลอโรพลาสต์ เช่น ระบบรับแสง คลอโรฟิลล์ Photosystem II, Photosystem I และกระบวนการตรึงคาร์บอนใน Calvin-Benson cycle
แต่การใช้แสงให้เกิดประโยชน์ไม่ได้ขึ้นอยู่กับแสงเพียงปัจจัยเดียว
ใบต้องอาศัยปัจจัยอื่นร่วมด้วย เช่น
- น้ำ
- คาร์บอนไดออกไซด์
- อุณหภูมิที่เหมาะสม
- การเปิดปิดของปากใบ
- ความพร้อมของเอนไซม์ในระบบตรึงคาร์บอน
- ความพร้อมของธาตุอาหาร
- ระบบรากที่ส่งน้ำและธาตุอาหารขึ้นมาเลี้ยงใบ
- ความสามารถในการระบายพลังงานส่วนเกินและซ่อมแซมระบบแสง
ถ้าแสงแรง แต่พืชยังมีน้ำพอ ปากใบทำงานได้ดี อุณหภูมิใบไม่สูงเกิน และระบบสังเคราะห์แสงยังใช้พลังงานได้ทัน แสงนั้นอาจยังเป็นประโยชน์
แต่ถ้าแสงแรงร่วมกับอากาศร้อน น้ำไม่พอ ปากใบปิด หรือระบบรากยังไม่พร้อม พืชอาจรับพลังงานแสงเข้ามามาก แต่ใช้พลังงานนั้นต่อได้ไม่เต็มที่
ตรงนี้เองที่แสงส่วนเกินเริ่มกลายเป็นความเครียดต่อใบ
2. Photoinhibition คืออะไร
Photoinhibition คือภาวะที่ประสิทธิภาพของระบบสังเคราะห์แสงลดลงจากการได้รับพลังงานแสงมากเกินกว่าระบบจะนำไปใช้หรือระบายออกได้ทัน
จุดที่มักเกี่ยวข้องมากคือระบบแสงที่ 2 หรือ Photosystem II (PSII) ซึ่งเป็นหนึ่งในศูนย์กลางสำคัญของปฏิกิริยาแสงในคลอโรพลาสต์
เมื่อระบบรับแสงได้รับพลังงานมากเกินไป พลังงานที่ส่งต่อไม่ทันอาจเพิ่มโอกาสการเกิด Reactive Oxygen Species หรือ ROS เช่น singlet oxygen, superoxide และ hydrogen peroxide
ROS เหล่านี้อาจเกี่ยวข้องกับความเสียหายของเม็ดสี คลอโรฟิลล์ เยื่อหุ้มเซลล์ โปรตีน และโครงสร้างภายในคลอโรพลาสต์ โดยเฉพาะโปรตีน D1 ซึ่งเป็นองค์ประกอบสำคัญของ Photosystem II
อย่างไรก็ตาม Photoinhibition ไม่ได้แปลว่าใบพืชเสียหายถาวรทันทีเสมอไป
ในระดับหนึ่ง ภาวะนี้อาจเกิดขึ้นชั่วคราว และพืชสามารถฟื้นตัวได้ หากความเครียดไม่รุนแรงเกินไป และระบบซ่อมแซมยังทำงานทัน
แต่ถ้าความเสียหายเกิดเร็วกว่าอัตราการซ่อมแซม โดยเฉพาะเมื่อแดดแรงร่วมกับความร้อน น้ำไม่พอ และปากใบปิดบ่อย ภาวะนี้อาจสะสมจนกลายเป็น Chronic photoinhibition หรือการยับยั้งการสังเคราะห์แสงแบบเรื้อรังได้
3. กลไกการหลีกเลี่ยงแสงทางกายภาพและสัณฐานวิทยา
ก่อนที่พืชจะต้องใช้กลไกระดับโมเลกุล ใบพืชมีด่านป้องกันระดับกายภาพที่ช่วยลดปริมาณแสงที่เข้าสู่ระบบสังเคราะห์แสงโดยตรง
กลไกกลุ่มนี้ไม่ได้ทำให้พืชหยุดรับแสง แต่ช่วยลดภาระของระบบใบเมื่อแสงแรงเกินไป
3.1 ผิวใบที่ช่วยสะท้อนแสง
พืชบางชนิดที่อยู่ในสภาพแสงจัดหรือแห้งแล้ง มักมีการปรับตัวทางสัณฐานวิทยา เช่น
- ชั้นคิวทิเคิลหนา
- แว็กซ์เคลือบผิวใบ
- ขนใบ หรือ Trichomes
- ผิวใบสีเทา สีเงิน หรือขาวนวล
โครงสร้างเหล่านี้ช่วยสะท้อนรังสีบางส่วนออกไป ลดการดูดซับพลังงานแสงและความร้อนเข้าสู่ใบ
ในแปลงปลูกหรือพื้นที่ปลูก เราอาจเห็นพืชบางชนิดมีใบสีซีด เทา หรือมีขนหนา ซึ่งไม่ได้เป็นอาการผิดปกติเสมอไป แต่อาจเป็นลักษณะการปรับตัวของพืชต่อสภาพแดดจัด
3.2 การปรับมุมใบเพื่อลดแสงตกกระทบ
พืชบางชนิดสามารถปรับทิศทางของใบให้หลบแสงได้ เช่น การเอียงใบให้ขนานกับทิศทางของแสงมากขึ้น เพื่อลดพื้นที่ที่แสงตกกระทบโดยตรง
กลไกนี้เรียกว่า Paraheliotropism
ในสภาพแดดจัด การปรับมุมใบช่วยลดทั้งภาระจากแสงและความร้อน โดยเฉพาะช่วงกลางวันที่ความเข้มแสงและอุณหภูมิสูง
ในพืชบางกลุ่ม อาจพบการม้วนใบหรือหุบใบเพื่อลดพื้นที่รับแสงและลดการสูญเสียน้ำร่วมด้วย
3.3 การเคลื่อนที่หลบแสงของคลอโรพลาสต์
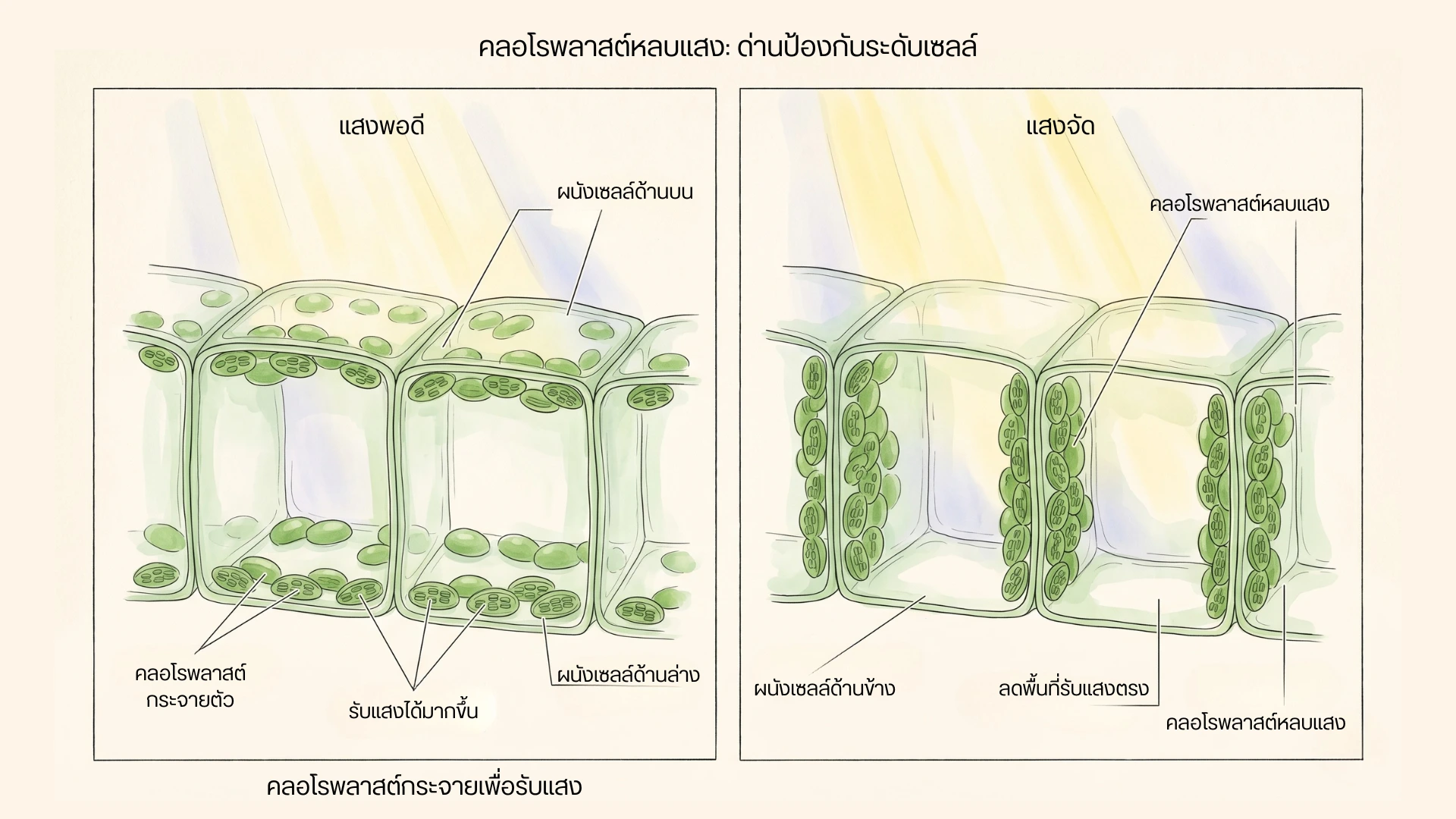 ภาพเปรียบเทียบตำแหน่งคลอโรพลาสต์ในเซลล์ใบระหว่างสภาพแสงพอดีกับสภาพแสงจัด โดยฝั่งแสงจัดคลอโรพลาสต์เคลื่อนไปเรียงตามผนังด้านข้างของเซลล์
ภาพเปรียบเทียบตำแหน่งคลอโรพลาสต์ในเซลล์ใบระหว่างสภาพแสงพอดีกับสภาพแสงจัด โดยฝั่งแสงจัดคลอโรพลาสต์เคลื่อนไปเรียงตามผนังด้านข้างของเซลล์
เมื่อแสงจัด คลอโรพลาสต์สามารถปรับตำแหน่งภายในเซลล์ เพื่อลดการรับแสงโดยตรงและลดภาระของระบบสังเคราะห์แสง
คลอโรพลาสต์ไม่ได้อยู่กับที่เสมอไป
ในสภาพแสงน้อย คลอโรพลาสต์มักจัดตำแหน่งเพื่อเพิ่มการรับแสง แต่เมื่อแสงจัดเกินไป คลอโรพลาสต์สามารถเคลื่อนที่ไปเรียงตัวบริเวณผนังเซลล์ด้านข้าง หรือ Anticlinal walls เพื่อลดพื้นที่รับแสงโดยตรง
กลไกนี้เรียกว่า Chloroplast avoidance movement
การเคลื่อนที่นี้เกี่ยวข้องกับการรับรู้แสงสีน้ำเงินผ่านตัวรับแสงกลุ่ม Phototropins และอาศัยระบบโครงร่างภายในเซลล์ เช่น Actin microfilaments
ในเชิงระบบ กลไกนี้เป็นเหมือนการปรับตำแหน่ง “แผงรับแสง” ภายในเซลล์ เพื่อช่วยลดภาระของระบบรับแสงเมื่อพืชเจอสภาพแสงจัด
4. Non-Photochemical Quenching: การระบายพลังงานส่วนเกินเป็นความร้อน
แม้พืชจะลดการรับแสงได้บางส่วน แต่พลังงานแสงจำนวนมากยังคงเข้าสู่ระบบรับแสงของใบ
ถ้าพลังงานนี้ถูกใช้ต่อในกระบวนการสังเคราะห์แสงไม่ทัน พืชต้องมีระบบระบายพลังงานส่วนเกินออกอย่างปลอดภัย
กลไกสำคัญคือ Non-Photochemical Quenching หรือ NPQ
NPQ คือกระบวนการที่พืชระบายพลังงานแสงส่วนเกินออกในรูปของความร้อน แทนที่จะปล่อยให้พลังงานนั้นไหลเข้าสู่ศูนย์กลางปฏิกิริยาและเพิ่มความเสี่ยงต่อความเสียหาย
กลไกนี้สำคัญมากในสภาพแสงเปลี่ยนแปลงระหว่างวัน เช่น แดดจัดสลับเมฆ หรือช่วงกลางวันที่ใบได้รับแสงแรงต่อเนื่อง
NPQ จึงทำหน้าที่เป็นระบบป้องกันระดับโมเลกุลที่ช่วยลดภาระของ Photosystem II และลดโอกาสการเกิด ROS จากพลังงานส่วนเกิน
5. Xanthophyll Cycle และบทบาทของ Zeaxanthin
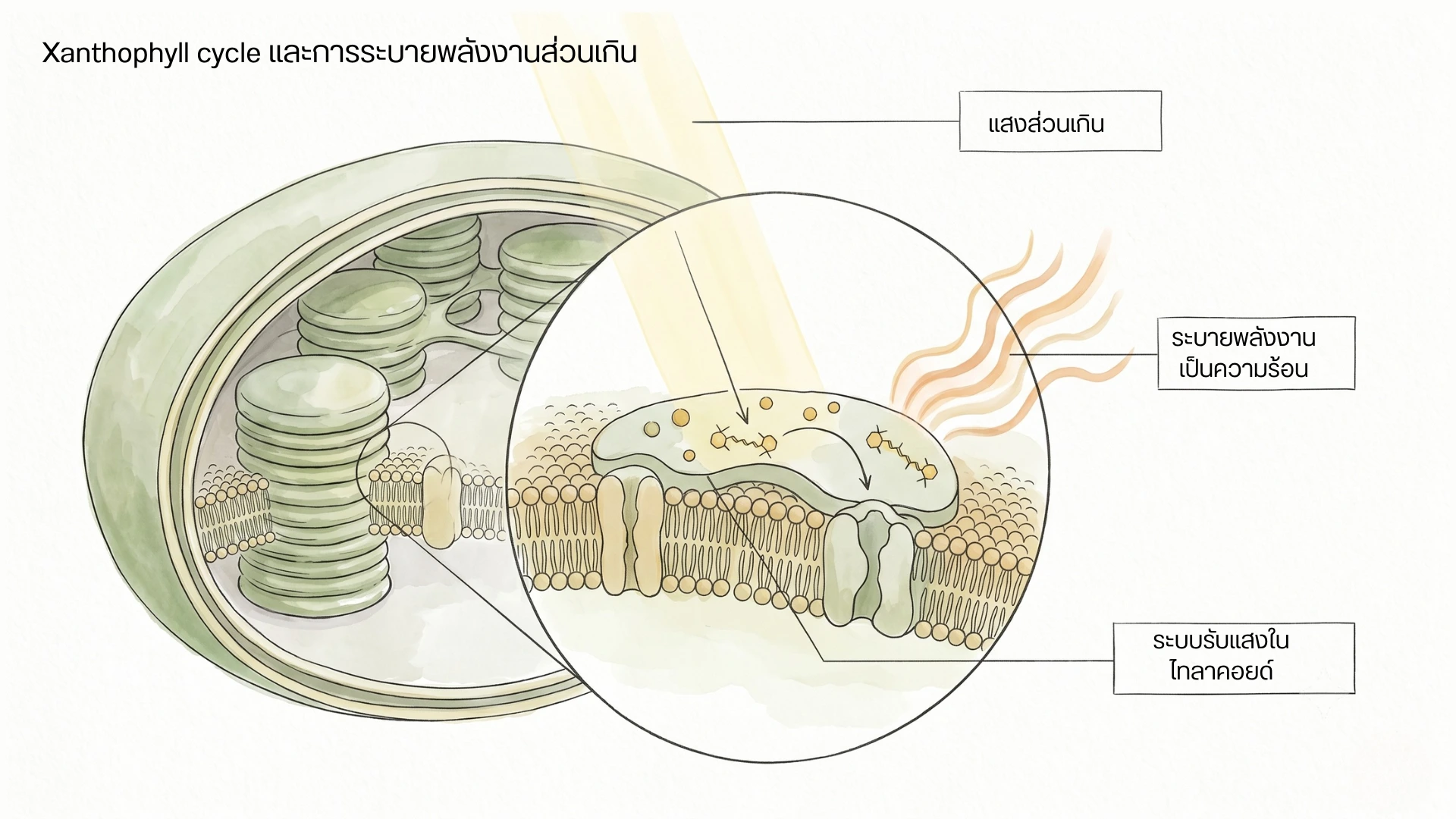 ภาพจำลองคลอโรพลาสต์และเยื่อไทลาคอยด์ แสดงพลังงานแสงเข้าสู่ระบบรับแสง เม็ดสีกลุ่มแซนโทฟิลล์ และการระบายพลังงานส่วนเกินออกเป็นความร้อน
ภาพจำลองคลอโรพลาสต์และเยื่อไทลาคอยด์ แสดงพลังงานแสงเข้าสู่ระบบรับแสง เม็ดสีกลุ่มแซนโทฟิลล์ และการระบายพลังงานส่วนเกินออกเป็นความร้อน
เมื่อพลังงานแสงมากเกินกว่าที่ใบใช้ได้ทัน พืชสามารถระบายพลังงานส่วนเกินบางส่วนออกเป็นความร้อนผ่านกลไก NPQ และ Xanthophyll cycle
หนึ่งในกลไกหลักที่เกี่ยวข้องกับ NPQ คือวัฏจักรแซนโทฟิลล์ หรือ Xanthophyll cycle
วัฏจักรนี้เกี่ยวข้องกับการเปลี่ยนรูปของรงควัตถุกลุ่มแคโรทีนอยด์ ได้แก่
- Violaxanthin
- Antheraxanthin
- Zeaxanthin
เมื่อใบได้รับแสงมาก การไหลของอิเล็กตรอนในปฏิกิริยาแสงทำให้ภายในลูเมนของไทลาคอยด์มีสภาพเป็นกรดมากขึ้น สภาพนี้กระตุ้นให้เกิดการเปลี่ยน Violaxanthin ไปเป็น Antheraxanthin และ Zeaxanthin
Zeaxanthin มีบทบาทสำคัญในการช่วยระบายพลังงานส่วนเกินออกเป็นความร้อน และทำงานร่วมกับองค์ประกอบอื่นของระบบรับแสง เช่น โปรตีน PsbS และ Antenna complex
เมื่อความเข้มแสงลดลง ระบบสามารถเปลี่ยน Zeaxanthin กลับไปเป็นรูปที่เหมาะกับการรับและส่งต่อพลังงานแสงเข้าสู่ระบบสังเคราะห์แสงได้มากขึ้น
จุดสำคัญคือ Xanthophyll cycle เป็นระบบที่ยืดหยุ่นและย้อนกลับได้ ช่วยให้ใบพืชปรับตัวต่อความเข้มแสงที่เปลี่ยนแปลงในแต่ละช่วงของวัน
6. ROS และระบบควบคุมอนุมูลอิสระ
หากพลังงานส่วนเกินระบายออกไม่ทัน ROS อาจเกิดขึ้นภายในคลอโรพลาสต์และส่วนอื่นของเซลล์
ROS หรือ Reactive Oxygen Species เป็นกลุ่มโมเลกุลออกซิเจนที่มีความไวต่อปฏิกิริยาเคมี เช่น
- Singlet oxygen
- Superoxide
- Hydrogen peroxide
ในระดับที่เหมาะสม ROS อาจเกี่ยวข้องกับการส่งสัญญาณภายในเซลล์ แต่เมื่อเกิดมากเกินไปหรือควบคุมไม่ได้ ROS อาจทำให้เกิดความเสียหายต่อเม็ดสี คลอโรฟิลล์ เยื่อหุ้มเซลล์ โปรตีน และส่วนประกอบต่าง ๆ ในคลอโรพลาสต์
พืชจึงมีระบบควบคุม ROS หลายกลุ่ม เช่น
- Carotenoids
- Zeaxanthin
- Flavonoids
- Anthocyanins
- Antioxidant enzymes บางกลุ่ม
Carotenoids และ Zeaxanthin มีบทบาททั้งในด้านการระบายพลังงานส่วนเกินและการช่วยลดผลกระทบจาก ROS ในบางบริบท
ส่วน Flavonoids และ Anthocyanins อาจสะสมบริเวณชั้นผิวใบ หรือ Epidermis และช่วยกรองรังสีบางช่วงคลื่น รวมถึงเกี่ยวข้องกับการตอบสนองต่อความเครียดจากแสงและ UV
อย่างไรก็ตาม ระบบ antioxidant ไม่ได้ทำงานแบบสารชนิดเดียวจบ แต่เป็นเครือข่ายของกลไกหลายระดับที่ทำงานร่วมกับสภาพน้ำ อุณหภูมิ และความพร้อมของเซลล์พืช
7. D1 Repair Cycle: การซ่อมแซม Photosystem II
แม้พืชจะมีระบบลดการรับแสง ระบายพลังงาน และควบคุม ROS แต่ในสภาพแสงจัด ความเสียหายต่อ Photosystem II ยังสามารถเกิดขึ้นได้
โปรตีน D1 เป็นองค์ประกอบสำคัญของ PSII และเป็นหนึ่งในส่วนที่มักได้รับผลกระทบจากภาวะ Photoinhibition
พืชจึงมีระบบซ่อมแซมที่เรียกว่า D1 repair cycle
โดยภาพรวม กระบวนการนี้ประกอบด้วย
- การตรวจจับหรือระบุโปรตีน D1 ที่เสียหาย
- การรื้อส่วนประกอบบางส่วนของ PSII
- การย่อยสลายโปรตีน D1 ที่เสียหาย
- การสังเคราะห์โปรตีน D1 ใหม่
- การประกอบกลับเข้าสู่ Photosystem II
วงจรนี้ช่วยให้ระบบสังเคราะห์แสงสามารถฟื้นตัวได้ หากอัตราความเสียหายไม่สูงเกินความสามารถในการซ่อมแซมของพืช
แต่ในสภาพแดดจัดต่อเนื่องร่วมกับความร้อนสูง น้ำไม่พอ หรือความเครียดอื่น ระบบซ่อมแซมอาจทำงานไม่ทัน ความเสียหายจึงอาจสะสมและส่งผลต่อประสิทธิภาพการสังเคราะห์แสง
8. Heat Stress, Water Stress และ Stomatal Closure
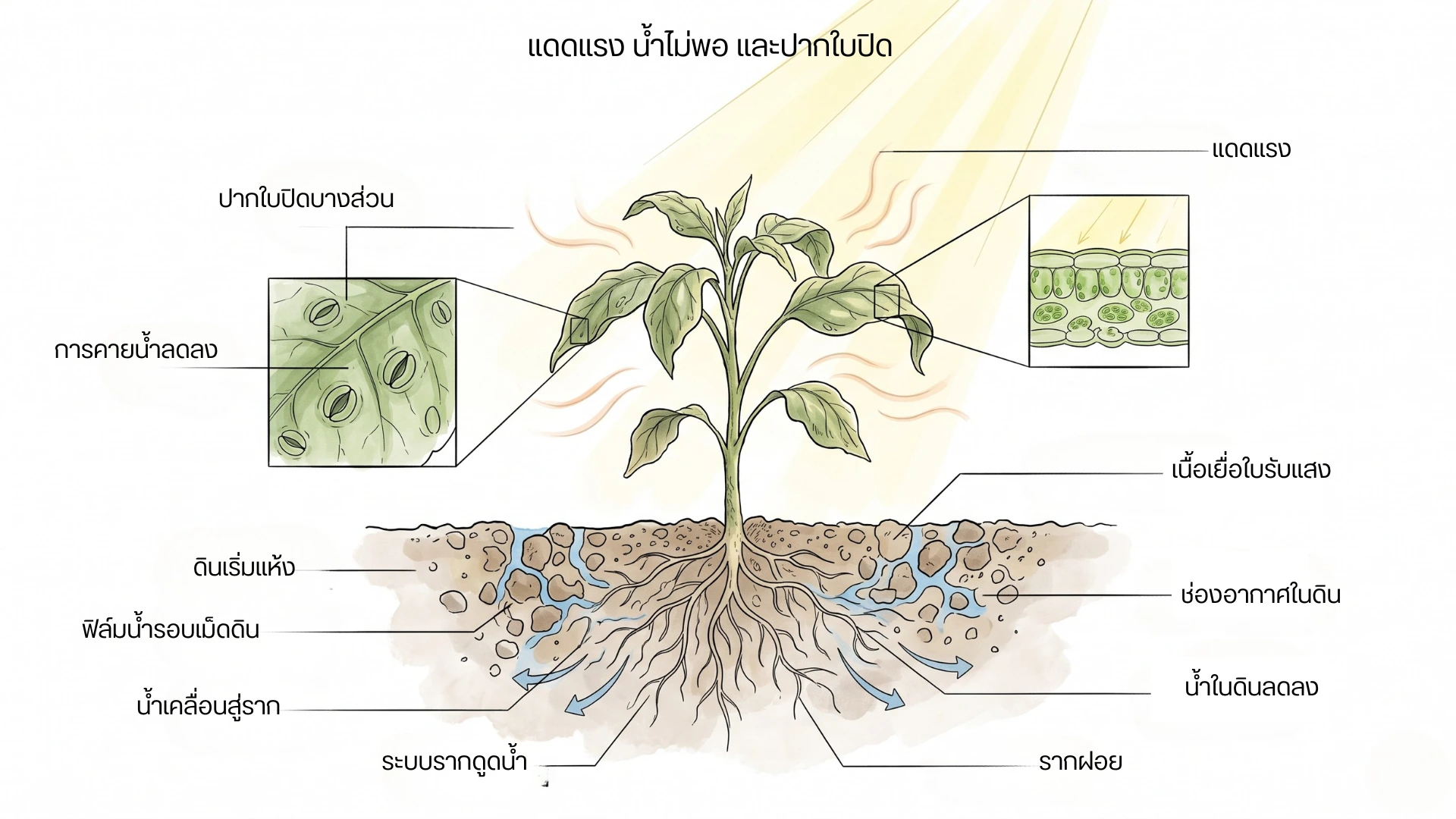 ภาพต้นพืชภายใต้แดดแรงและความร้อน มีใบลู่เล็กน้อย ระบบรากในดินที่มีน้ำลดลง และภาพขยายปากใบกับเนื้อเยื่อใบ เพื่ออธิบายความสัมพันธ์ระหว่างแสงจัด น้ำไม่พอ และการปิดปากใบ
ภาพต้นพืชภายใต้แดดแรงและความร้อน มีใบลู่เล็กน้อย ระบบรากในดินที่มีน้ำลดลง และภาพขยายปากใบกับเนื้อเยื่อใบ เพื่ออธิบายความสัมพันธ์ระหว่างแสงจัด น้ำไม่พอ และการปิดปากใบ
เมื่อแดดแรงเกิดร่วมกับน้ำไม่พอและอุณหภูมิสูง ปากใบอาจปิดเพื่อลดการสูญเสียน้ำ แต่การปิดปากใบก็ทำให้ CO₂ เข้าสู่ใบได้น้อยลง และอาจเพิ่มภาระต่อระบบสังเคราะห์แสง
ในแปลงปลูกจริง ความเครียดจากแสงมักเกิดร่วมกับความเครียดจากความร้อนและน้ำ
เมื่อพืชขาดน้ำ ปากใบมักปิดเพื่อลดการสูญเสียน้ำ การปิดปากใบช่วยรักษาน้ำในต้น แต่ทำให้ CO₂ เข้าสู่ใบได้น้อยลง
เมื่อ CO₂ ลดลง กระบวนการตรึงคาร์บอนใน Calvin-Benson cycle อาจถูกจำกัด ในขณะที่ใบยังได้รับพลังงานแสงต่อเนื่อง
สภาพนี้ทำให้เกิดความไม่สมดุลระหว่างพลังงานที่รับเข้ามา กับความสามารถในการนำพลังงานนั้นไปใช้สร้างสารอินทรีย์
นอกจากนี้ การคายน้ำยังช่วยระบายความร้อนของใบ เมื่อการคายน้ำลดลง อุณหภูมิใบอาจสูงขึ้น และเพิ่มภาระต่อเยื่อหุ้มเซลล์ โปรตีน และเอนไซม์ในระบบสังเคราะห์แสง
นี่คือเหตุผลที่แดดแรงในวันที่น้ำพอ กับแดดแรงในวันที่น้ำไม่พอ อาจส่งผลต่อพืชไม่เหมือนกัน
แสงจึงควรถูกอ่านร่วมกับน้ำ อุณหภูมิ ปากใบ และสภาพรากเสมอ
9. Rubisco Activase และข้อจำกัดทางชีวเคมีเมื่อใบอุณหภูมิสูง
นอกจากปฏิกิริยาแสงแล้ว การสังเคราะห์ด้วยแสงยังต้องพึ่งพากระบวนการตรึงคาร์บอนไดออกไซด์
Rubisco เป็นเอนไซม์หลักในกระบวนการตรึง CO₂ แต่การทำงานของ Rubisco ต้องอาศัย Rubisco activase หรือ Rca ช่วยรักษาสถานะการทำงานของเอนไซม์ให้พร้อมใช้งาน
ในพืชหลายชนิด Rca ค่อนข้างอ่อนไหวต่อความร้อน เมื่ออุณหภูมิใบสูงขึ้นมาก ความสามารถของ Rca อาจลดลง ทำให้ Rubisco ทำงานได้ไม่เต็มที่ และการตรึง CO₂ ลดลง
จุดนี้ทำให้เห็นว่า ในสภาพแดดแรงและอากาศร้อน ปัญหาไม่ได้อยู่ที่แสงเพียงอย่างเดียว แต่เกี่ยวข้องกับความสามารถของระบบชีวเคมีภายในใบในการใช้พลังงานแสงต่อด้วย
อย่างไรก็ตาม ความทนทานของ Rca แตกต่างกันตามชนิดพืช สายพันธุ์ และสภาพแวดล้อมเดิมของพืช จึงไม่ควรใช้ตัวเลขอุณหภูมิเดียวไปสรุปกับพืชทุกชนิด
สำหรับคนปลูกพืช ประเด็นนี้ช่วยอธิบายได้ว่า ทำไมบางช่วงที่แดดจัดมาก พืชอาจไม่ได้สร้างอาหารเพิ่มขึ้นตามปริมาณแสง แต่กลับชะงัก เหี่ยว โทรม หรือฟื้นตัวช้าลง โดยเฉพาะเมื่อมีความร้อนและน้ำไม่พอร่วมด้วย
10. ปัจจัยสนับสนุนความพร้อมของระบบสังเคราะห์แสง: ธาตุอาหาร ฮอร์โมน และจุลินทรีย์
กลไกการป้องกันแสงส่วนเกินไม่ได้เกิดขึ้นเฉพาะในคลอโรพลาสต์เท่านั้น
แม้ NPQ, Xanthophyll cycle, ROS scavenging และ D1 repair cycle จะเป็นกลไกสำคัญภายในใบ แต่ความสามารถของพืชในการรับมือกับแสงจัดยังขึ้นอยู่กับความพร้อมของระบบทั้งต้นด้วย
โดยเฉพาะ 3 กลุ่มปัจจัยที่เกี่ยวข้องกันอย่างใกล้ชิด ได้แก่
- ธาตุอาหาร
- ฮอร์โมนพืช
- จุลินทรีย์ที่อยู่ร่วมกับระบบราก
ปัจจัยเหล่านี้ไม่ได้ทำหน้าที่เป็นเกราะกันแดดโดยตรง แต่ช่วยสนับสนุนให้ระบบสังเคราะห์แสง ระบบน้ำ ระบบราก และการซ่อมแซมภายในเซลล์มีความพร้อมมากขึ้น ภายใต้เงื่อนไขที่เหมาะสม
10.1 ธาตุอาหาร: โครงสร้างและตัวร่วมของระบบสังเคราะห์แสง
ธาตุอาหารหลายชนิดเกี่ยวข้องกับความสามารถของใบในการรับ ใช้ ระบาย และซ่อมแซมผลกระทบจากพลังงานแสง
ไนโตรเจน (N) และแมกนีเซียม (Mg) ไนโตรเจนเป็นองค์ประกอบของกรดอะมิโน โปรตีน เอนไซม์ และโปรตีนจำนวนมากในระบบสังเคราะห์แสง รวมถึง Rubisco ซึ่งเป็นเอนไซม์หลักในกระบวนการตรึงคาร์บอนไดออกไซด์
แมกนีเซียมเป็นอะตอมกลางของโมเลกุลคลอโรฟิลล์ และยังเกี่ยวข้องกับการทำงานของเอนไซม์หลายกลุ่มที่ต้องใช้ ATP รวมถึงระบบที่ช่วยให้ Rubisco อยู่ในสถานะพร้อมทำงาน
ดังนั้น เมื่อ N หรือ Mg ไม่เพียงพอ ความสามารถในการสร้างคลอโรฟิลล์ โปรตีน และเอนไซม์ที่เกี่ยวข้องกับการสังเคราะห์แสงอาจลดลง ทำให้ใบมีความพร้อมต่อแสงลดลงตามไปด้วย
เหล็ก (Fe) เหล็กเกี่ยวข้องกับระบบถ่ายทอดอิเล็กตรอนในคลอโรพลาสต์ เช่น Cytochromes และ Ferredoxin รวมถึงกระบวนการบางส่วนที่เกี่ยวข้องกับการสร้างคลอโรฟิลล์
เมื่อพืชขาดเหล็ก ใบอ่อนมักมีอาการซีดหรือเหลือง เนื่องจากระบบสร้างคลอโรฟิลล์และการทำงานของคลอโรพลาสต์ถูกรบกวน
แมงกานีส (Mn), แคลเซียม (Ca) และคลอรีน (Cl) ธาตุกลุ่มนี้เกี่ยวข้องกับ Oxygen-Evolving Complex หรือ OEC ใน Photosystem II ซึ่งเป็นระบบที่ทำหน้าที่แยกน้ำและส่งอิเล็กตรอนเข้าสู่ปฏิกิริยาแสง
ในเชิงกลไก OEC มีความสำคัญต่อการรักษาการไหลของอิเล็กตรอนใน PSII หากระบบนี้ทำงานผิดปกติ ภาระต่อศูนย์กลางปฏิกิริยาอาจเพิ่มขึ้น และอาจเกี่ยวข้องกับความเสี่ยงต่อความเสียหายของระบบแสง
อย่างไรก็ตาม ไม่ควรสรุปว่าการให้ Mn, Ca หรือ Cl เพิ่ม จะป้องกัน Photoinhibition ได้โดยตรง เพราะความเสียหายจากแสงส่วนเกินขึ้นอยู่กับหลายปัจจัยร่วมกัน ทั้งน้ำ อุณหภูมิ ปากใบ สภาพราก และความสามารถในการซ่อมแซมของพืช
ฟอสฟอรัส (P) ฟอสฟอรัสเป็นองค์ประกอบของ ATP, กรดนิวคลีอิก และกระบวนการถ่ายทอดพลังงานหลายส่วนในเซลล์
ในบริบทของความเครียดจากแสง P เกี่ยวข้องกับความพร้อมของพลังงานและกระบวนการสร้างโปรตีนใหม่ ซึ่งมีความสำคัญต่อการซ่อมแซมระบบที่เสียหาย เช่น การสร้างโปรตีน D1 ใหม่ใน Photosystem II
โพแทสเซียม (K) โพแทสเซียมเกี่ยวข้องกับการควบคุมแรงดันเต่งของเซลล์คุม และมีบทบาทสำคัญต่อการเปิดปิดปากใบ
เมื่อปากใบทำงานสมดุล พืชสามารถรับ CO₂ เพื่อใช้ในการตรึงคาร์บอน และคายน้ำเพื่อช่วยระบายความร้อนของใบได้ดีขึ้น
แต่ถ้าน้ำไม่พอ แม้ K จะเกี่ยวข้องกับการควบคุมปากใบ พืชก็ยังอาจต้องปิดปากใบเพื่อลดการสูญเสียน้ำอยู่ดี
ดังนั้น ธาตุอาหารควรถูกมองเป็นส่วนหนึ่งของความพร้อมของระบบสังเคราะห์แสง ไม่ใช่ปัจจัยเดี่ยวที่ทำให้พืชทนแดดจัดได้เสมอ
10.2 ฮอร์โมนพืช: ระบบสื่อสารเมื่อพืชเผชิญความเครียด
ฮอร์โมนพืชทำหน้าที่เป็นระบบสื่อสารภายในต้น เมื่อพืชเผชิญความเครียดจากแสง ความร้อน หรือการขาดน้ำ
Abscisic Acid (ABA) ABA เป็นฮอร์โมนสำคัญที่เกี่ยวข้องกับการตอบสนองต่อความแห้งแล้ง
เมื่อรากหรือใบรับรู้ว่าน้ำไม่เพียงพอ พืชอาจเพิ่มการสร้าง ABA ซึ่งส่งสัญญาณไปยังเซลล์คุม ทำให้ปากใบปิดเพื่อลดการสูญเสียน้ำ
การปิดปากใบช่วยรักษาน้ำในต้น แต่ก็มีผลข้างเคียงคือ CO₂ เข้าสู่ใบได้น้อยลง เมื่อ CO₂ ลดลง กระบวนการตรึงคาร์บอนอาจถูกจำกัด ในขณะที่ใบยังได้รับพลังงานแสงต่อเนื่อง
ดังนั้น ABA มีบทบาทสำคัญต่อการเอาตัวรอดจากภาวะขาดน้ำ แต่ในอีกด้านหนึ่งก็อาจทำให้เกิดความไม่สมดุลระหว่างพลังงานแสงที่รับเข้ามากับพลังงานที่ใช้ได้ในกระบวนการสังเคราะห์แสง
Cytokinins ไซโทไคนินเป็นฮอร์โมนที่เกี่ยวข้องกับการเจริญของเซลล์ ความสมบูรณ์ของใบ และการชะลอความเสื่อมของเนื้อเยื่อในบางบริบท
มีรายงานในบางการศึกษาว่า Cytokinins อาจเกี่ยวข้องกับการรักษากิจกรรมของระบบสังเคราะห์แสงภายใต้สภาวะเครียดบางแบบ แต่ผลลัพธ์ขึ้นอยู่กับชนิดพืช ระยะการเจริญ และระดับความรุนแรงของความเครียด
Salicylic Acid (SA) และ Brassinosteroids (BRs) SA และ BRs เป็นกลุ่มสารควบคุมการเจริญและการตอบสนองต่อความเครียดที่มีรายงานว่าเกี่ยวข้องกับระบบ antioxidant การทำงานของเอนไซม์ และการลดผลกระทบจากความเครียดบางชนิด
ในบางการศึกษา การให้ SA หรือ BRs จากภายนอกมีความสัมพันธ์กับการฟื้นตัวของระบบสังเคราะห์แสงภายใต้ความเครียด เช่น ความร้อน ภัยแล้ง หรือพิษจากโลหะหนัก
อย่างไรก็ตาม การแปลผลเรื่องฮอร์โมนพืชต้องระวังมาก เพราะผลลัพธ์ขึ้นอยู่กับความเข้มข้น วิธีใช้ ชนิดพืช อายุพืช และสภาพแวดล้อม ไม่ควรนำไปสรุปเป็นสูตรใช้งานทั่วไปโดยตรง
10.3 จุลินทรีย์ที่มีประโยชน์: การสนับสนุนจากระบบรากและไรโซสเฟียร์
ในธรรมชาติ พืชไม่ได้รับมือกับความเครียดเพียงลำพัง
บริเวณรอบราก หรือ Rhizosphere เป็นพื้นที่ที่รากพืช จุลินทรีย์ น้ำ ธาตุอาหาร และอินทรียวัตถุมีปฏิสัมพันธ์กันอย่างต่อเนื่อง
จุลินทรีย์บางกลุ่มอาจช่วยสนับสนุนความพร้อมของพืชต่อความเครียดจากแสงและความร้อน ผ่านการช่วยระบบราก น้ำ และธาตุอาหาร มากกว่าการป้องกันแสงโดยตรง
Arbuscular Mycorrhizal Fungi (AMF) ราไมคอร์ไรซาบางกลุ่มสามารถอยู่ร่วมกับรากพืช และช่วยขยายพื้นที่ดูดซับน้ำและธาตุอาหาร โดยเฉพาะธาตุที่เคลื่อนที่ในดินได้จำกัด เช่น ฟอสฟอรัส รวมถึงธาตุรองและจุลธาตุบางชนิด
เมื่อระบบรากเข้าถึงน้ำและธาตุอาหารได้ดีขึ้น พืชอาจรักษาสถานะน้ำและความพร้อมของระบบสังเคราะห์แสงได้ดีขึ้นในบางสภาพแวดล้อม
Plant Growth-Promoting Rhizobacteria (PGPR) แบคทีเรียส่งเสริมการเจริญเติบโตของพืชบางกลุ่มในบริเวณราก อาจช่วยพืชผ่านหลายกลไก เช่น
- การสร้างสารคล้ายฮอร์โมนบางชนิด
- การตรึงไนโตรเจนในบางระบบ
- การละลายฟอสเฟต
- การสร้าง Siderophores ที่ช่วยเกี่ยวข้องกับการนำเหล็กเข้าสู่ระบบ
- การส่งเสริมการพัฒนารากในบางบริบท
กลไกเหล่านี้อาจช่วยให้พืชมีระบบรากและโภชนาการที่ดีขึ้น ซึ่งส่งผลทางอ้อมต่อความสามารถในการรับมือกับความเครียดจากแสง ความร้อน และน้ำไม่พอ
จุลินทรีย์ทนเครียดบางกลุ่ม มีการศึกษาบางส่วนเกี่ยวกับจุลินทรีย์ที่ช่วยพืชรับมือกับความเครียดเฉพาะ เช่น ความเค็ม ความแห้งแล้ง หรืออุณหภูมิสูง
บางรายงานพบว่าจุลินทรีย์เหล่านี้อาจช่วยรักษาประสิทธิภาพการสังเคราะห์แสงหรือการทำงานของเอนไซม์บางส่วนในพืชบางชนิดได้
อย่างไรก็ตาม ผลลัพธ์ของจุลินทรีย์ขึ้นอยู่กับชนิดพืช สายพันธุ์จุลินทรีย์ สภาพดิน ความชื้น อุณหภูมิ และการจัดการในระบบปลูก จึงควรสื่อสารอย่างระมัดระวังว่าเป็นการสนับสนุนภายใต้เงื่อนไขที่เหมาะสม ไม่ใช่ผลที่เกิดขึ้นแน่นอนในทุกกรณี
10.4 มองแบบระบบ: ใบไม่ได้ทำงานแยกจากรากและดิน
เมื่อพืชเจอแดดจัด ใบเป็นพื้นที่ที่รับผลกระทบชัดที่สุด แต่ความสามารถในการรับมือไม่ได้อยู่ที่ใบเพียงอย่างเดียว
ใบต้องอาศัยน้ำจากราก รากต้องอาศัยดินหรือวัสดุปลูกที่มีความชื้นเหมาะสม ธาตุอาหารต้องอยู่ในรูปที่พืชเข้าถึงได้ ปากใบต้องปรับตัวให้สมดุลระหว่างการรับ CO₂ และการลดการสูญเสียน้ำ ระบบสังเคราะห์แสงต้องมีเอนไซม์ เม็ดสี และโครงสร้างที่พร้อม จุลินทรีย์ในไรโซสเฟียร์อาจช่วยสนับสนุนระบบรากและโภชนาการในบางบริบท
ดังนั้น การจัดการพืชให้รับมือกับแดดจัดไม่ได้เริ่มที่ใบอย่างเดียว แต่เริ่มตั้งแต่ระบบราก น้ำ ดิน วัสดุปลูก ธาตุอาหาร และสภาพแวดล้อมรอบต้น
ในมุมของ Green Fineness นี่คือเหตุผลที่การอ่านอาการใบควรถูกเชื่อมกลับไปยังระบบทั้งต้นเสมอ
11. ความหมายเชิงการปลูกพืช
สำหรับคนปลูกพืช บทเรียนสำคัญของเรื่องนี้คือ แสงมากไม่ได้มีผลเหมือนกันในทุกสภาพ
แสงจัดอาจเป็นประโยชน์เมื่อพืชมีน้ำเพียงพอ ระบบรากสมบูรณ์ ปากใบทำงานได้ดี และอุณหภูมิใบไม่สูงเกินไป
แต่ในสภาพที่น้ำไม่พอ รากยังไม่แข็งแรง วัสดุปลูกแห้งเร็ว หรืออากาศร้อนจัด แสงจัดอาจกลายเป็นภาระต่อใบได้ง่ายขึ้น
การสังเกตภาคสนามจึงควรดูปัจจัยร่วม เช่น
- แสงแรงช่วงเวลาใดของวัน
- ใบฟื้นตัวหลังแดดลดลงหรือไม่
- วัสดุปลูกแห้งเร็วหรือไม่
- พืชเพิ่งย้ายปลูกหรืออยู่ในช่วงตั้งตัวหรือไม่
- มีลมร้อนหรืออุณหภูมิสะสมในพื้นที่ปลูกหรือไม่
- ใบมีอาการซีด ไหม้ ม้วน ลู่ หรือชะงักต่อเนื่องหรือไม่
- ระบบรากยังทำงานได้ดีหรือมีปัญหา
- น้ำและธาตุอาหารอยู่ในระดับเหมาะสมหรือไม่
จุดสำคัญคือ ไม่ควรอ่านใบจากอาการเดียว
ใบซีดไม่ได้แปลว่าขาดไนโตรเจนเสมอ ใบไหม้ไม่ได้แปลว่าแดดแรงเพียงอย่างเดียว ใบลู่ไม่ได้แปลว่าพืชกำลังจะตายเสมอไป และแดดแรงไม่ได้แปลว่าพืชจะสร้างอาหารได้มากขึ้นเสมอ
อาการที่เห็นบนใบควรถูกอ่านร่วมกับระบบน้ำ อุณหภูมิ ปากใบ ราก ธาตุอาหาร และสภาพแวดล้อมทั้งหมด
สรุป
แสงเป็นพลังงานสำคัญของพืช แต่เมื่อแสงมากเกินกว่าที่ใบจะใช้ได้ทัน พลังงานแสงส่วนเกินอาจกลายเป็นภาระต่อระบบสังเคราะห์แสง
พืชจึงมีกลไกป้องกันหลายระดับ ตั้งแต่การลดแสงที่ตกกระทบใบ การเคลื่อนที่ของคลอโรพลาสต์ การระบายพลังงานผ่าน NPQ และ Xanthophyll cycle การควบคุม ROS ไปจนถึงการซ่อมแซม Photosystem II ผ่าน D1 repair cycle
ในสภาพปลูกจริง ความเครียดจากแสงมักเกิดร่วมกับความร้อน น้ำไม่พอ ปากใบปิด และข้อจำกัดของระบบรากหรือเอนไซม์ภายในใบ เช่น Rubisco activase
ธาตุอาหาร ฮอร์โมนพืช และจุลินทรีย์ในไรโซสเฟียร์ อาจมีบทบาทสนับสนุนความพร้อมของระบบสังเคราะห์แสงภายใต้เงื่อนไขที่เหมาะสม แต่ไม่ควรถูกมองเป็นปัจจัยเดี่ยวที่ทำให้พืชทนแดดจัดได้โดยตรง
สำหรับคนปลูกพืช ประเด็นสำคัญจึงไม่ใช่การถามว่า แดดดีหรือแดดไม่ดี
แต่คือการดูว่า แสง น้ำ อุณหภูมิ ปากใบ ราก ธาตุอาหาร และระบบซ่อมแซมของพืช ยังทำงานร่วมกันได้สมดุลหรือไม่
เพราะสำหรับพืชแล้ว การอยู่กลางแดดไม่ใช่แค่การรับพลังงาน
แต่คือการรักษาสมดุลของทั้งระบบใบ ให้สามารถใช้พลังงานแสงได้โดยไม่ทำร้ายเครื่องจักรสังเคราะห์แสงของตัวเอง
Internal Link Suggestions ถ้าแสงน้อย พืชยังสร้างอาหารได้ไหม ใบเริ่มทำงาน: จากใบอ่อน สู่ระบบสร้างอาหารของพืช ปากใบ น้ำ และอากาศร้อน ใบ พืชไม่ได้ใช้เพียงไนโตรเจน ระบบลำเลียงของพืช: Xylem / Phloem ในช่วงต้นกล้า ไรโซสเฟียร์: พื้นที่ที่รากเริ่มเชื่อมกับดิน จุลินทรีย์ในดินทำอะไรบ้าง Reference Note
เอกสารอ้างอิง
-
Taiz, L., Zeiger, E., Møller, I. M., & Murphy, A. Plant Physiology and Development. Sinauer Associates / Oxford University Press.
-
Hopkins, W. G., & Hüner, N. P. A. Introduction to Plant Physiology.
-
Demmig-Adams, B., & Adams, W. W. งานวิชาการด้าน Photoprotection, Non-photochemical quenching และ Xanthophyll cycle ในพืช.
-
งานวิจัยเรื่อง The Dynamics of Energy Dissipation and Xanthophyll Conversion in Arabidopsis. Frontiers.
-
งานวิจัยเรื่อง Mechanism of Nonphotochemical Quenching in Green Plants. Biochemistry / ACS Publications.
-
Ehleringer, J. R., Björkman, O., & Mooney, H. A. งานวิจัยเกี่ยวกับ leaf pubescence, surface reflectance และการปรับตัวของใบพืชต่อแสงจัด.
-
Gorton, H. L., Williams, W. E., & Vogelmann, T. C. งานวิจัยเกี่ยวกับ chloroplast movement และการตอบสนองของคลอโรพลาสต์ต่อแสง.
-
Haupt, W., & Scheuerlein, R. งานวิจัยเกี่ยวกับกลไกการเคลื่อนที่ของคลอโรพลาสต์และการหลีกเลี่ยงแสงในพืช.
-
Buchanan, B. B., Gruissem, W., & Jones, R. L. Biochemistry & Molecular Biology of Plants. American Society of Plant Biologists.
-
Lambers, H., Chapin, F. S., & Pons, T. L. Plant Physiological Ecology. Springer.
-
Marschner, P. Marschner’s Mineral Nutrition of Higher Plants. Academic Press.


